The morphogenetic field that determines pattern in early embryogenesis may be written in liquid crystal phase alignments through electrodyamic patterning processes Dr. Mae-Wan Ho
Liquid crystals are a fourth state of matter between liquid and solid [1]. There are many kinds of liquid crystals, each with distinctive chemical structure and properties. The hall mark of liquid crystals is long-range orientation order imposed by molecular anisotropy (e.g., long thin molecules) and electrical dipole interactions. This long-range orientation order defines their special optical, mechanical electrical and magnetic properties. It also makes liquid crystals responsive to electric and magnetic fields, finding many applications in liquid crystal devices (LCDs) such as TV and computer displays, digital watches, calculators, etc.
Liquid crystals are piezoelectric, or flexoelectric [2], in that they interconvert mechanical energy and electrical polarization. They are similarly sensitive to temperature and pH. In other words, liquid crystals integrate and interconvert all forms of energy within the organism, which is why they are ideal for making organisms (see my book [3] The Rainbow and the Worm, The Physics of Organisms, ISIS publication) and may well also provide the ideal active substrate for pattern formation (as suggested in Chapter 15 of the book). In the next report, I shall present recent evidence in support of the idea that living organisms are animated and organised by electronic induction and electromagnetic intercommunication [4] (Electronic Induction Animates the Cell, SiS 52).
There is already a substantial body of work on pattern formation in liquid crystals iin response to electromagnetic fields. Electroconvection (flow induced by electromagnetic gradients) in nematic (rod-like) liquid crystals has been intensively studied over the past two decades, and provides a rich variety of pattern-formation phenomena [5]. Stripes and cells and more chaotic patterns can form in thin sheets of nematic liquid crystals according to the field conditions. The dynamic patterns in Figure 1 were obtained in a 1 x 1 cm slab about 100 mm thick.

Figure 1 Patterns in a layer of nematic liquid crystals exposed to different electric and magnetic fields viewed under polarized light microscopy, assembled from [5]
In addition to the non-equilibrium ‘dissipative’ structures that depend on continuous input of energy, liquid crystals can undergo equilibrium transitions driven by an electric field, governed by a minimization of free energy to give equilibrium structures [2]. Equilibrium liquid crystalline structures on the surface of cells and embryos are reminiscent of the ‘morphogenetic field’ that appears to direct pattern formation, discovered by the pioneers of embryology such as Alexander Gurwitsch, (1876-1954) in Russia, Hans Spemann (1869-1941) in Germany, and Paul Weiss (1898-1989) in the United States [6], but has remained mysterious to this day.
At the start of development, the embryo is totipotent, in other words, all parts of it have the potential to become many structures, or any structure of the adult organisms. In the course of early development, however, a hierarchy of determination events occur, so that different parts of the embryo become more and more restricted in their developmental potential, but with as yet no sign of the structures they are determined to form at some time in the future. The determined state could only be demonstrated by transplant or grafting experiments.
A small part removed from an embryo before determination and transplanted to a different location, or grated to another embryo, will develop in harmony with its surroundings. If the same experiment is carried out after determination, however, the graft will develop the structure it was determined to be, irrespective of its surroundings. Thus, the graft may develop into a limb on the back of the host, for example.
The paradoxical nature of the determined state is that the determinative influences not only possesses field-like characteristics, but is also material and transplantable. This has spurred generations of developmental biologists to hunt, in vain to this very day, for specific chemical ‘morphogens’, despite the fact that vast numbers of genes and gene products have been identified and mapped. One could claim that it is the special combination of genes expressed that makes specific determined states, but there is nothing in the genes and gene products that could account for the field-like effect (see [7] Genes Don't Generate Body Patterns, SiS 52).
In the late 1980s, pioneering biophysicist Lynn Trainor (1921-2008), a good friend, now deceased, then at University of Toronto in Canada, and his graduate student John Totafurno decided to re-examine the result of classic experiments in which limb-buds, literally precursors of limbs on the body of developing salamander embryos, were transplanted upside-down, left-right interchanged, or variously rotated to another stump. When these limb-buds eventually developed, extra limbs were often induced in strange configurations (see Figure 2).
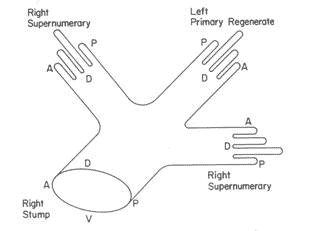
Figure 2 Result of grafting a left limb bud onto a right stump in salamander; A, P, D, V represent anterior, posterior, dorsal, and ventral respectively
Totafurno and Trainor successfully interpreted all the results in terms of a nonlinear 2-dimensional vector field on a cylinder, in which the oriented field lines need to reconnect (to become continuous) as smoothly as possible [8, 9]. The smooth connections between anterior, posterior, dorsal and ventral fields of the stump and the graft are evident in the resulting regenerated limb (Figure 2), even though it takes two extra hands to do the trick.
I have suggested that the vector field might be literally written in the orientation of biological liquid crystals [3]; perhaps especially those associated with the cell membrane and the underlying cortical actin-myosin network.
In support of this idea, early Drosophila embryos, briefly exposed to static magnetic fields at the time when crucial pattern determination processes are known to be taking place, developed characteristic ‘twisted’ deformities in the larvae that hatch some 24 hours later [10, 11]. The normal segmented pattern (Figure 3 a) is transformed into various helical configurations (Figure 3b-h).
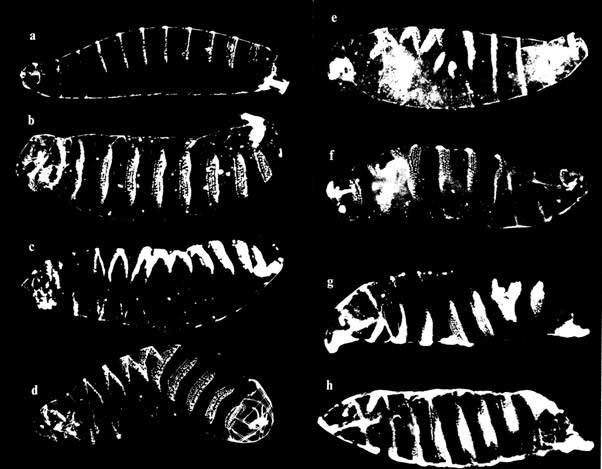
Figure 3 Various helical transformations (b-h) of the normal segmental pattern (a) in Drosophila larvae that have been briefly exposed to a weak static magnetic field during early embryogenesis
The significance of the results is that they were obtained by exposure to a static magnetic field, which must be acting on electrodynamic processes. Furthermore, the energy involved is several orders of magnitude below the ‘thermal threshold’ (of a system without coherence). Consequently, there can be no effects unless there is a high degree of coherence among the molecules involved in the pattern determination processes, and the most likely candidates for these molecules are liquid crystals. The morphogenetic ‘field’ effect is evident in the global nature of the transformations, the most perfect being the complete helix of larva h in Figure 3.
The morphogenetic field that determines pattern in early embryogenesis may be written in liquid crystal phase alignments through electrodyamic patterning processes. This hypothesis can now be tested directly with the sophisticated polarized light imaging techniques available.
Article first published 05/10/11
Got something to say about this page? Comment
There are 2 comments on this article so far. Add your comment above.
Warren Brodey M.D. Comment left 6th October 2011 07:07:57
Kindly send a list of references.
This is an important
Thanks for making it available to your mem
warren
michael Comment left 6th October 2011 19:07:41
what do you think the ramifactions of this are for electromagnetic radiationand it's effects on the developing foetus?